高棘龙
高棘龙(学名Acrocanthosaurus),又名高脊龙、多脊龙或阿克罗肯龙,意为「有高棘的蜥蜴」,是肉食龙下目异特龙超科鲨齿龙科的一个属,生活在白垩纪早期到中期北美洲的美国,约1.2亿至1.08亿年前。如同大部分恐龙的属,高棘龙只有单一种,阿托卡高棘龙。它们的化石发现于美国的奥克拉荷马州与德克萨斯州、怀饿明州等州,在马里兰州也则发现了属于高棘龙的牙齿。最大高棘龙的标本可长11.8米,体重达到7吨。

高棘龙 - 叙述
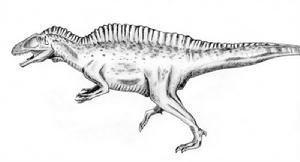
高棘龙与人类的体型相比图册高棘龙是一种大型的双足肉食性恐龙。如同名称所显示,牠们的脊椎有很多部份都有高大的神经突,极可能支撑者由肌肉所构成的隆脊,从颈部延伸到背部、臀部。高棘龙是最大型的兽脚亚目恐龙之一,身长接近12公尺,重量可达约2.4公吨。在德州所发现的大型兽脚类足迹,可能是由高棘龙所留下,但没有相关骨骸可以直接证实。
近年发现的化石增添了许多生理结构的特征,可进一步研究高棘龙的脑部结构与前肢功能。然而,高棘龙的演化关系仍存在者争论,有些科学家将牠们归类于异特龙科,其它科学家则归之于鲨齿龙科。高棘龙是牠们所处生态系统的最大型兽脚类恐龙,并可能是顶级掠食者,以大型蜥脚类与鸟脚类恐龙为食。
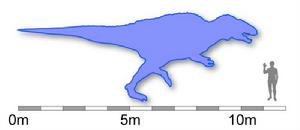
高棘龙的体型略小于牠们的大型近亲南方巨兽龙,但仍是最大型的兽脚类恐龙之一。目前已知最大的个体为11公尺长,重量估计为2,400公斤。头颅骨的长度接近1.3公尺。
如同大部分其它的兽脚类恐龙,高棘龙的头颅骨长、低矮、狭窄。眶前孔相当大,约占头部的1/4长,2/3高度,可减轻头部的重量。上颚骨两侧与鼻骨上侧的表面,并不像南方巨兽龙与鲨齿龙那样多起伏。鼻骨上有长而低矮的棱脊,从鼻孔开始,沿者口鼻部的两侧,直到眼睛处的泪骨。所有异特龙超科的头部都具有这种棱脊。不像异特龙,高棘龙的眼睛前方的泪骨并没有角冠。牠们的泪骨与眶后骨愈合,在眼睛上方形成一个厚眉脊,这点与鲨齿龙科、没有亲缘关系的阿贝力龙科相同。上颚两侧各有19颗锯齿状、弯曲的牙齿,但下颚的牙齿数量未明。高棘龙的牙齿比鲨齿龙的还长,而且没有鲨齿龙科特有的牙齿纹路。下颚的齿骨前缘呈方形,如同南方巨兽龙;齿骨的上下宽度窄,而颚部后段骨头的上下宽度非常宽。高棘龙与南方巨兽龙的下颚上隅骨侧边都有宽厚、水平方向的棱脊。

阿托卡高棘龙想象图图册高棘龙最明显的特征是在牠们的高大的神经突,从颈部延伸到背部、臀部及尾巴前段,长度可以达脊椎长度的2.5倍。其它在背部有着高大背棘的恐龙,有些的背棘比高棘龙的更巨大。例如,生存于非洲的棘龙,棘龙的背棘约为2米高,约是所连接的脊椎骨的11倍长。与棘龙的背棘支撑者皮肤不同,高棘龙的背棘较低矮,可能作为腱壮肌肉的附着处,类似现代的美洲野牛,并在背部形成一个高而厚的隆脊。背棘的功能仍然未明,可能跟动物之间的沟通、储存脂肪或控制体温有关。所有颈椎与背椎的两侧都有明显的侧腔,而尾椎的侧腔较小。这个特征较为类似鲨齿龙科,而非异特龙科。
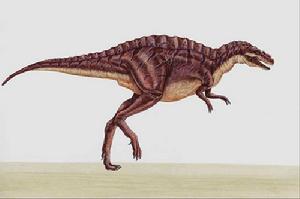
除了脊椎以外,高棘龙具有典型的异特龙超科骨骸。高棘龙是种二足恐龙,尾巴长而重,可平衡头部与身体的重量,将重心保持在臀部。与异特龙相比,高棘龙的前肢较短、较粗壮,但其它方面很类似,手部都有三根手指,上有指爪。与小型的善奔恐龙不同,牠们的股骨长于胫骨与跖骨,这显示高棘龙不善于快速奔跑。相较于异特龙,高棘龙的后肢骨头较为粗壮。如同其它兽脚类恐龙,高棘龙的脚掌具有四根脚趾,第一趾小于其它小趾,无法接触地面。
高棘龙 - 分类系统
现代的亲缘分支分析法将高棘龙归类于坚尾龙类的异特龙超科中,但在这个超科中的位置却被有争议。异特龙超科的特征是鼻骨与泪骨上方有一对棱脊,颈椎上有高大的神经突,以及其它特征。高棘龙最初与异特龙被分类为异特龙科中,后期的一些研究亦支持这个说法。很多其它的科学家却将牠与鲨齿龙及南方巨兽龙分类在鲨齿龙科内。
如同大部分其它的大型兽脚类恐龙,高棘龙最初被发现时只有零散的化石,使牠们的分类经常改变。J. Willis Stovall与Wann Langston, Jr.最初将高棘龙分类于腔躯龙科(Antrodemidae),是异特龙科的同义词。在1956年,阿尔弗雷德•罗默(Alfred Sherwood Romer)将高棘龙改列为斑龙科,该科过去曾被当作「未分类物种集中地」。在60年代,其它科学家认为牠们脊椎上的长神经突,代表者与棘龙的亲缘关系。将高棘龙归类于棘龙科的分类法,持续到80年代,并出现在当时的半科学文献当中。
在1884年,英格兰的早白垩纪地层发现了高大的神经突,曾经被认为非常类似高棘龙。这些骨头最初被归类于顶棘龙,顶棘龙是种英国的兽脚类恐龙,仅发现牙齿。在1988年,葛瑞格利•保罗(Gregory S. Paul)在1988年将这些神经突命名为高棘龙的第二个种,长棘高棘龙(Acrocanthosaurus altispinax)。保罗的归类使得至少一位科学家主张顶棘龙是高棘龙的一个异名。在1991年,长棘高棘龙被改建立为独立的比克尔斯棘龙,不属于高棘龙与顶棘龙。
如果高棘龙属于异特龙亚科,牠们的最近亲应为侏罗纪的异特龙与食蜥王龙。那些将高棘龙归类于鲨齿龙科的亲缘分支分类法研究,常将高棘龙置于比非洲鲨齿龙与南美洲的南方巨兽龙更为原始的位置。发现于英格兰的新猎龙,经常被认为是种更基础的鲨齿龙科动物。新猎龙意味者鲨齿龙科起源于欧洲,而后散布到南方的冈瓦那大陆。如果高棘龙属于鲨齿龙科,意味者鲨齿龙科也散布到北美洲。目前已知的鲨齿龙科都生存于白垩纪的早期到中期。
高棘龙 - 发现与命名
已架设的高棘龙骨骸图册高棘龙的名字是从其高大的神经突而来,ακρα/akra在古希腊文的意思是「高的」,ακανθα/akantha意为「棘」或「刺」,而σαυρος/sauros意为「蜥蜴」。高棘龙的唯一种阿托卡高棘龙(Acrocanthosaurus atokensis),是按其发现地方,俄克拉何马州的阿托卡县来命名。高棘龙的名称是由美国古生物学家J. Willis Stovall及Wann Langston, Jr.于1950年命名的。Langston曾在他于1947年的未公布硕士论文中,提到了Acracanthus atokaensis这个名称,但在正式公布时,则改为阿托卡高棘龙。

高棘龙的完模标本
(编号OMNH 10146)及副模标本(编号OMNH 10147),包含两个部份骨骼及一少片的头颅骨,是同时于1950年命名,并从美国俄克拉何马州的鹿角组发现。在90年代,两副较完整的骨骼最近亦被发现。第一副标本(编号SMU 74646)是个部份骨骸,约为70%完整,大部份的头颅骨遗失,是从得克萨斯州的双子山地层发现,现时保存于沃斯堡科学历史博物馆内。另一个更完整的骨骼(编号NCSM 14345)通常昵称为「法兰」,是由私人收藏家发现于鹿角组,经过南达科塔州的黑山地质研究院 处理过后,目前保存于北卡罗莱纳州罗利市的北卡罗莱纳州自然科学博物馆内。这个标本保有目前唯一的完整高棘龙头颅骨与前肢,也是目前最大型的标本。高棘龙的副模标本与「法兰」的大小接近,显示牠们的体型大致接近,而编号SMU 74646则明显较小。很多其它的碎片及独立骨头在得克萨斯州北部、俄克拉何马州及甚至亚利桑那州。
俄克拉何马州与德州以外的地区,可能已发现高棘龙的化石,但较不完整。一个发现于亚利桑那州南部的牙齿,可能属于高棘龙,在该地区的蜥脚类化石也发现了符合高棘龙的齿痕。发现于马里兰州的阿伦德尔地层的数颗牙齿,与高棘龙的牙齿几乎一致,可能为高棘龙的东部代表。美国西部的许多地层组发现的牙齿与骨头,曾经被归类于高棘龙,但大部分是归类错误。
高棘龙 - 古生物学
前肢功能
高棘龙的左手部模型图册如同大部分其它的兽脚类恐龙,高棘龙的前肢无法接触地面,因此没有行走的功能,而是在猎食时发生作用。「法兰」的完整前肢,有助于研究高棘龙前肢的功能与活动范围。一个研究检验了骨头与其相连骨头的表面,以计算关节能有多少活动范围,而不会关节脱位。如同许多现存初龙类,许多手部关节的骨头并没有完全相吻合,显示这些关节有一定的软骨。这个研究并指出当高棘龙休息时,前肢会从肩膀下垂,肱骨微向后摆,手肘弯曲,指爪朝内。

人类相比,高棘龙的肩膀的转动范围很小。牠们的手臂无法做出360°的旋转幅度,但可后摆至离垂直面约109°,所以肱骨可以后摆至斜微上方。手臂可以前转至超过垂直面约24°。手臂往无法垂直地往下摆,但可外展至超于水平面约9°。与人类相比,手肘的活动范围也很小,大约只有57°的转动幅度。 手臂无法完全地伸直,也无法大幅弯曲,而肱骨不能做出90°弯曲。桡骨与尺骨互相固定,所以无法如人类的前臂,做出往内侧或外侧旋转的动作。
个腕骨之间没有准确地相接合,显示手腕有大量的软骨,可使手腕坚韧。当手指向外扳时,能几乎碰触到手腕。手指向内弯曲时,第一指能与第二指平行,第三指只能内弯。第一指的指爪最大,而且总是往内弯曲。第二指爪也总是弯曲的,而最小的第三指爪则可往内侧、外侧摆动。
除了计算前肢关节的活动范围以外,这个研究还提出了高棘龙的猎食习性假设。牠们的前肢的前摆幅度不大,无法勾抓猎物的背部。由于前肢无法在捕捉猎物 开始时派上用场,高棘龙可能主要使用嘴部来猎食。此外,前肢无法将猎物大幅拉近。只有当嘴部咬住猎物,强壮的前肢才能将猎物拉近、紧紧抓住,防止猎物逃脱。当猎物企图逃脱时,高棘龙可用弯曲的第一、第二指爪刺住猎物的身体。手指的大幅外扳幅度是种演化适应,可让高棘龙不用在关节脱落的风险下,抓住挣扎的猎物。一旦猎物被紧抓到身体旁,高棘龙便可用颚部吞咬牠们。另一种可能的猎食方式则是高棘龙以颚部咬住猎物,并用前肢不断地往自己拉,用指爪划出大型的伤口。
脑部与内耳结构
阿托卡高棘龙的头部图册2005年,科学家们以X射线断层成像分析高棘龙完模标本脑壳的内部空间,而制作了一个高棘龙的颅腔模型。在牠们生前,这些颅内空间除了装有脑部,应该还有脑膜与脑脊液。这个颅腔模型可用来判定脑部及脑神经的一般特征,也可与其它的兽脚类颅腔模型做比较。高棘龙的脑部类似大部分的兽脚类恐龙,最为类似异特龙超科。牠们的脑部看似鲨齿龙及南方巨兽龙,多于异特龙或中华盗龙,这支持了高棘龙属于鲨齿龙科的假说。
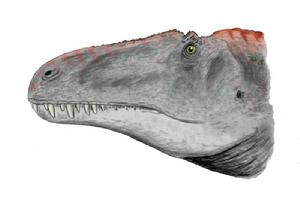
高棘龙的脑部形状稍微呈S形,大脑半球并不甚扩张,近似鳄鱼多于鸟类。大部分的非虚骨龙类兽脚类恐龙都具有这项特征。高棘龙的嗅球很大,呈球根状,可见牠们的嗅觉很好。耳朵半规管的重建显示头部是保持在与水平面呈25°夹角。这是透过将模型调校至侧半规管与地表成平行,显示这种动物有者处于警戒中的步态。
2005年,科学家们以X射线断层成像分析高棘龙完模标本脑壳的内部空间,而制作了一个高棘龙的颅腔模型。在牠们生前,这些颅内空间除了装有脑部,应该还有脑膜与脑脊液。这个颅腔模型可用来判定脑部及脑神经的一般特征,也可与其它的兽脚类颅腔模型做比较。高棘龙的脑部类似大部分的兽脚类恐龙,最为类似异特龙超科。牠们的脑部看似鲨齿龙及南方巨兽龙,多于异特龙或中华盗龙,这支持了高棘龙属于鲨齿龙科的假说。
高棘龙的脑部形状稍微呈S形,大脑半球并不甚扩张,近似鳄鱼多于鸟类。大部分的非虚骨龙类兽脚类恐龙都具有这项特征。高棘龙的嗅球很大,呈球根状,可见牠们的嗅觉很好。耳朵半规管的重建显示头部是保持在与水平面呈25°夹角。这是透过将模型调校至侧半规管与地表成平行,显示这种动物有者处于警戒中的步态。
高棘龙 - 古生态学 阿托卡高棘龙想象图图册高棘龙的化石发现于德州北部的双子山组与俄克拉何马州南部的鹿角组。这些地层还没有经过放射性同位素计年,但科学家们利用生物地层学来估计它们的年代。根据菊石化石的变化,德州的玫瑰组地层位于早白垩纪阿普第阶与艾伯塔阶的交界,该地层发现过高棘龙的可能足迹,而且位于双子山组的上层。这显示双子山组都属于阿普第阶,而阿普第阶的年代为1亿2500万年前到1亿1200万年前。鹿角组的化石还包含恐爪龙与腱龙,两者的化石也发现于蒙大拿州的Cloverly组,Cloverly组经过测量后,被发现年代为阿普第阶至阿尔布阶,这显示鹿角组的年代应该相同。因此,高棘龙极可能生存于1亿2500万年前到1亿年前间。
在这个时期,双子山组与鹿角组是大片泛滥平原,位于陆缘海附近。在数百万年之后,这片海洋向北扩张,形成西部内陆海道,并将北美洲一分为二,持续到几乎整个晚白垩纪。玫瑰谷地层是个沿岸环境,高棘龙的可能足迹应该是留在古代海岸边的泥泞平地上。由于高棘龙是种大型掠食者,牠们被推测生存于大范围的地区,居住于许多不同的环境。高棘龙的猎食对象可能有:蜥脚类的帕拉克西龙(Paluxysaurus)、体积庞大的波塞东龙、以及大型鸟脚类恐龙如腱龙。[40]恐爪龙也生存于同一地区,牠们是种小型兽脚类恐龙,身长约3公尺,极可能与高棘龙有少许的竞争。
高棘龙足迹
在德克萨斯州中部的玫瑰谷地层,保有许多恐龙的足迹,包含大型、三趾兽脚亚目的足迹。其中最著名的足迹化石发现于恐龙谷州立公园的巴拉斯河附近,目前正在纽约美国自然历史博物馆展示中,[29]该地附近也发现了数个足迹化石。 [30][31]由于没有化石被发现,这不可能界定这些足迹是属于什么动物。但长久以来,科学家认为这些足迹可能与高棘龙有关。 [32]一个2001年的研究将玫瑰谷地层的足迹与不同的大型兽脚类恐龙的脚掌相比,但无法明确地归类于任何一属。但该研究发现这个足迹的形状与大小类似高棘龙。因为玫瑰谷地层与鹿角组、双子山组的地理位置与地质年代接近,而这些地区的唯一大型兽脚类恐龙为高棘龙,该研究推论这些足迹最可能由高棘龙所留下。
这个著名的兽脚亚目足迹属于数个个体,与多达12只蜥脚下目恐龙向着同一方向前进。有些兽脚亚目足迹是在蜥脚下目足迹之上,显示它们是在之后形成的,这显示有一小群高棘龙在追踪一群蜥脚下目恐龙。 [29]这个有趣的理论看似合理,但是却很难证实,而有其他解释。例如,在蜥脚类恐龙经过后,多只个别的兽脚类恐龙在不同时间经过同一方向,仿佛好像是在追踪猎物一般。同样的,也可以说蜥脚下目是个别的前进,而非整群的移动。 [34]当一批兽脚类足迹横越其中一批蜥脚类足迹时,缺乏一个脚印,这被视为该恐龙展开攻击的证据。 [35]但是,其他科学家对这解释提出质疑,因为该批蜥脚类足迹并未改变步态,如果有任何大型掠食者位在该蜥脚类恐龙身边,该恐龙应该会改变其姿势。
高棘龙古生态学
高棘龙的化石发现于德州北部的双子山组与奥克拉荷马州南部的鹿角组。这些地层还没有经过放射性同位素计年,但科学家们利用生物地层学来估计它们的年代。根据菊石化石的变化,德州的玫瑰组地层位于早白垩纪阿普第阶与阿尔比阶的交界,该地层发现过高棘龙的可能足迹,而且位于双子山组的上层。这显示双子山组都属于阿普第阶,而阿普第阶的年代为1亿2500万年前到1亿1200万年前。 [36]鹿角组的化石还包含恐爪龙与腱龙,两者的化石也发现于蒙大拿州的Cloverly组,Cloverly组经过测量后,被发现年代为阿普第阶至阿尔布阶,这显示鹿角组的年代应该相同。 [37]因此,高棘龙极可能生存于1亿2500万年前到1亿年前间。
在这个时期,双子山组与鹿角组是大片泛滥平原,位于陆缘海附近。在数百万年之后,这片海洋向北扩张,形成西部内陆海道,并将北美洲一分为二,持续到几乎整个晚白垩纪。玫瑰谷地层是个沿岸环境,高棘龙的可能足迹应该是留在古代海岸边的泥泞平地上。由于高棘龙是种大型掠食者,它们被推测生存于大范围的地区,居住于许多不同的环境。 [33]高棘龙的猎食对象可能有:蜥脚类的帕拉克西龙(Paluxysaurus)、[38]体积庞大的波塞东龙、[39]以及大型鸟脚类恐龙如腱龙。 [40]恐爪龙也生存于同一地区,它们是种小型兽脚类恐龙,身长约3公尺,极可能与高棘龙有少许的竞争。
高棘龙动画角色
古代王者恐龙王中的70只恐龙之一
普通型
高棘龙高棘龙(初次登场第15集)
属性:火
绝招:爆炎璧攻、爆炎大炮
在世界出现的地方:日本的酒店温泉
强化型
高棘龙(强化型)被强化后的高棘龙
高棘龙(强化型)(初次登场第16集)
属性:火
绝招:爆炎璧攻、爆炎大炮
在世界出现的地方:原本是亚法岛,后来是日本的三条市
