玛君龙
玛君龙(属名:Majungasaurus)别名玛君颅龙、玛宗格厚头龙又译玛宗格龙,意为“马任加蜥蜴”,是兽脚亚目阿贝力龙科的一属,生存于白垩纪末的马达加斯加,约7000万年前到6500万年前。目前仅有一个已确认种,凹齿玛君龙(M. crenatissimus)。玛君龙一度被认为是种厚头龙下目恐龙,玛君颅龙现被视为玛君龙的次异名。

中文名:玛君龙
别名:玛君颅龙、玛宗格厚头龙
英文名字:Majunga lizard/Lizard from Majunga
拉丁学名:Majungatholus
外文名:Majungasaurus crenatissimus
界:动物界
纲:蜥形纲
科:阿贝力龙科
属:玛君龙属
种:玛君龙
亚目:兽脚亚目
分布区域:马达加斯加
门:脊索动物门
目:蜥臀目
别称:玛君颅龙
时代:白垩纪
玛君龙基本资料
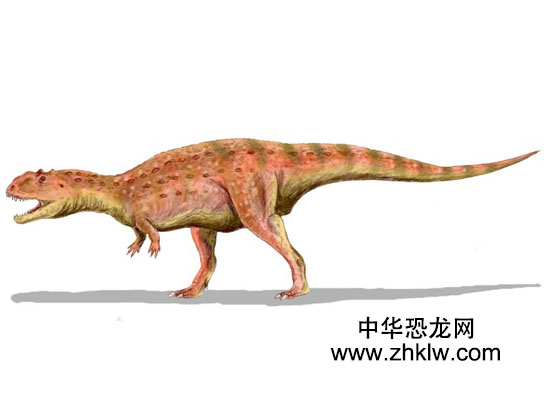
玛君龙如同其他阿贝力龙科恐龙,玛君龙是种二足掠食动物,拥有短口鼻部。虽然目前并不清楚前肢的 完整样貌,它们的前肢非常短,而后肢较长、较粗壮。玛君龙可从以下特征与其他阿贝力龙科恐龙分辨出来,例如:较厚的头颅骨、口鼻部上方的表面不平较厚骨头、以及头顶上的一根圆形角状物;这个角状物起初曾被误认为是厚头龙类的颅顶。玛君龙的上颌与下颌牙齿比大部分阿贝力龙科恐龙还多。 玛君龙目前已发现数个保存良好的头颅骨与大量的骨骸,并在近期成为南半球的兽脚亚目恐龙中最广受研究的一种。玛君龙似乎与印度的阿贝力龙科恐龙关系较近,而离南美洲与非洲的阿贝力龙科恐龙较远,这点在生物地理学上有重要的意义。对于玛君龙来说,它们最血腥的事就是同类相食。除了捕猎食草恐龙外,玛君龙还会自相残杀,捕食同类,真是一种可怕的恐龙啊!玛君龙目前已发现数个保存良好的头颅骨与大量的完整骨骸,是南半球的肉食恐龙中最广受研究的一种,玛君龙是该生态系统的顶级掠食者,主要猎食蜥脚下目恐龙,例如掠食龙;玛君龙也目前唯一有直接证据显示同类相食现象的恐龙。
玛君龙体形特征
大型恐龙,体长8米在目前已知的 752 种恐龙中,体形排在第 227位。与玛君龙 体形接近的恐龙,有艾伯塔龙,食肉牛龙,日本龙,楯甲龙,阿贝力龙,崇高龙等。
玛君龙是种中等体型的兽脚亚目恐龙,身长约6到7米,这数据包含尾巴在内。大型玛君龙个体的破碎化石显示有些成年体身长超过8米。科学家们估计成年的玛君龙平均体重超过1,100公斤,但其中最大型的个体可能超过这个数据。玛君龙的近亲食肉牛龙身长8到9米,重量经估计为1,500公斤。与大部分的兽脚亚目恐龙相比,玛君龙的头颅骨相当著名,而且类似其他阿贝力龙科恐龙的头颅。如同其他阿贝力龙科的头颅,玛君龙的头颅长度与高玛君龙体型度比例较短,但没有食肉牛龙那样短。大型成年个体的头颅骨长度约60到70厘米。前上颌骨使得口鼻部前端非常钝,这是阿贝力龙科恐龙的典型特征。然而,与其他阿贝力龙科恐龙的头颅骨相比,玛君龙的头颅骨宽度较宽。所有阿贝力龙科恐龙的头颅骨都有粗糙不平的表面,玛君龙也不例外。玛君龙的鼻骨特别厚,并且互相固定,鼻骨的下半部有个低鼻脊。头顶的固定额骨有个明显的半球形角状物。在玛君龙存活的时候,这些结构可能由某种结构所覆盖,可能由角质所构成。电脑断层扫描显示这些鼻部结构与额角拥有中空空间,可能用来减轻重量。玛君龙的牙齿有短齿冠,这是典型的阿贝力龙科特征;而玛君龙的上颌骨与齿骨分别有17颗牙齿,是阿贝力龙科当中最多的,除了皱褶龙以外。 玛君龙的颅后骨骼极度类似食肉牛龙与奥卡龙的颅后骨骼,这两个属是其他曾发现完整骨骸的阿贝力龙科恐龙。玛君龙是二足恐龙,拥有长尾巴,以平玛君龙衡头部与胸部,使得重心位在臀部。虽然颈椎拥有许多空间以减轻这些部位的重量,但还是有加大肌肉附着在上面,而肋骨互相连错、加强,所以颈部还是很结实。颈部肋骨有骨化肌腱附着,如同食肉牛龙。不同的是,玛君龙的颈部肋骨侧边有长凹处,以渐轻重量。综合以上特征,玛君龙的颈部强壮、充满肌肉。
肱骨是前肢唯一被叙述的部份,肱骨短而弯曲,极度类似食肉牛龙与奥卡龙的肱骨;这显示玛君龙可能拥有非常类似的短前肢与四指。如同其他阿贝力龙科恐龙,玛君龙的前肢使身体相比,短而结实。玛君龙的胫骨比食肉牛龙的胫骨还要结实,膝盖部位有明显的突出。距骨与跟骨互相固定者,脚部有3根具有功能的脚趾,而最小的第一趾并未接触到地面上。
玛君龙食性特征
科学家们提出玛君龙与其他阿贝力龙科恐龙的独特头颅形状,显示出与其它兽脚类恐龙不同掠食习性。大多数兽脚类恐龙有长、低矮、狭窄的头颅骨,阿贝力龙科恐龙的头颅骨较高且宽,长度通常较短。其他兽脚类恐龙的狭窄头颅骨可承受强壮咬合时的垂直压力,但不适合承受扭转力量。与现代哺乳类掠食者相比,大多数兽脚类恐龙可能使用类似犬科动物的方式猎食,使用长而狭窄的口鼻部多次咬伤猎物,使猎物变虚弱。
玛君龙是它们所处环境的最大型掠食者,而该地区同一时期的最大型草食性动物是蜥脚类恐龙,例如掠食龙。科学家们认为玛君龙,或者其他的阿贝力龙科恐龙,特别以蜥脚类恐龙作为猎食对象。它们头部与颈部的增强,以适应咬合与咬住的攻击方式,可能对于巨大、强壮的蜥脚类恐龙特别有效。相较于其他兽脚类恐龙较修长的后肢,玛君龙短而结实的后肢,可支持这个假设。虽然玛君龙的奔跑速度无法赶上相同大小的兽脚类恐龙时,它们仍可以追上移动缓慢的蜥脚类恐龙。玛君龙的结实后肢骨头显示它们拥有非常强壮的后腿,而后腿长度较短,可降低玛君龙的重心位置。因此玛君龙牺牲速度,以增加力量。在掠食龙骨头上发现的玛君龙牙齿痕迹,确认它们至少以这些蜥脚类恐龙为食,但不确定玛君龙是猎食它们,或是在它们死后吞食它们的尸体。
阿贝力龙科恐龙,尤其是玛君龙,可能改采用类似现代猫科动物的方式猎食,使用短而宽广的口鼻部咬住猎物紧紧不放,直到它们被制服。玛君龙的口鼻部较其他阿贝力龙科恐龙还宽广,而且它们的其他生理特征也支持者这个猎食方式假设。玛君龙的颈部较壮,且拥有结实的颈椎、交错的颈部肋骨、骨化的肌腱、以及附着在脊椎骨与头颅后方的强化肌肉。这些强化肌肉可使玛君龙的猎物挣扎时,玛君龙仍能保持头部的稳定性。阿贝力龙科恐龙的头颅骨有许多区域,有皮肤经过矿化作用而成的骨头,形成凹凸不平的表面。阿贝力龙科的鼻骨加厚且固定住,可增加强度,这种特征在玛君龙身上特别明显。另一方面,玛君龙的下颌两侧各有一个大型洞孔,这特征可见于其他角鼻龙类恐龙;特定关节间有滑液关节,可允许下颌的大幅度变形扩张,但不能达到蛇类的扩张程度。这些下颌特征可防止玛君龙嘴中的猎物挣扎时,下颌遭到破裂。下颌的前部牙齿比其他牙齿还要结实,可提供咬合时的支撑点;而玛君龙的低齿冠牙齿可防止猎物挣扎时,将牙齿摇断。最后,不像异特龙与大部分兽脚类恐龙,它们的牙齿呈前后侧弯曲;而阿贝力龙科恐龙的牙齿,例如玛君龙,它们的牙齿前侧弯曲,而后侧较直。这种牙齿结构可用来将猎物切碎,而非将猎物托在咬合的地方。
玛君龙分类
玛君龙被分类于兽脚亚目的阿贝力龙科演化支,而阿贝力龙科与西北阿根廷龙科共同组成阿贝力龙超科,阿贝力龙超科则是角鼻龙下目的一个分支。阿贝力龙科恐龙的特征是:高头颅骨、钝口鼻部、面部骨头的凹凸不平表面(与鲨齿龙科的趋同演化结果)、缩短的前肢(与暴龙科的趋同演化结果)、以及结实的后肢。
如同许多的恐龙科,阿贝力龙科内部成员的演化关系非常混淆。数个亲缘分支分类法研究显示在系统发生学上,玛君龙与南美洲的食肉牛龙有接近亲缘关系, 而其他属则无法确定演化位置。一个最近的研究,使用最完整的资讯,不将玛君龙与印度的胜王龙、印度龙分类于同一演化支,而是与南美洲的食肉牛龙、肌肉龙、爆诞龙、奥卡龙、阿贝力龙、以及非洲的皱褶龙分类于同一演化支。这个研究使得阿贝力龙科可能分别在西冈瓦纳大陆与东冈瓦纳大陆形成个别的次演化支[1]。需要更进一步的发现与研究才能协助了解阿贝力龙科与玛君龙的详细系统发生学。
发现与命名
在1896年,法国古生物学家Charles Depéret叙述了第一个来自于马达加斯加西北部的兽脚亚目化石。这些化石是由一个法国陆军军官在贝齐布卡河(Betsiboka River)沿岸发现,包含两颗牙齿、一个指爪、以及一些脊椎骨,目前存放在里昂第一大学。Depéret将这些化石分类于斑龙属的一种,凹齿斑龙(megalosaurus crenatissimus);斑龙属在当时是个“未分类物种集中地”,包含许多大型兽脚类,但这些物种彼此之间并没有关联。种名crenatissimus衍化自拉丁语,crenatus意为“凹槽”,而issimus意为“最”,意指它们牙齿前后的大量锯齿状边缘。Depéret稍后将它们重新归类于北美洲的伤龙,这是另一个所知有限的属[13]。玛君龙的模式标本,额骨的角曾被误认为是厚头龙类的颅顶。目前存放于法国国立自然史博物馆 玛君龙的下颌。位于法国国立自然史博物馆 在马达加斯加西北部马哈赞加省发现的许多破碎化石,在过去100多年来经由法国籍人员发现,其中许多目前存放在巴黎国家自然历史博物馆[1]。在1955年,René Lavocat在发现第一个标本的同一地区梅法拉诺组,发现了一个兽脚亚目的齿骨与牙齿。这些牙齿符合Depéret首次叙述的标本,但大幅弯曲的颌部骨头与斑龙、伤龙有极大不同。Lavocat将它们重新命名为玛君龙(Majungasaurus),属名由马哈赞加省的旧名马任加省(Majunga Province)以及希腊文的蜥蜴所构成,并将这个颌部骨头列为模式标本(编号MNHN.MAJ 1),存放在巴黎国家自然历史博物馆。在1979年,汉斯·戴尔特·苏伊士(Hans-Dieter Sues)与菲利普·塔丘特(Philippe Taquet)将玛君龙第四号标本(编号MNHN.MAJ 4)的圆顶头颅碎片,叙述成一个新的厚头龙类恐龙,名为玛君颅龙(Majungatholus atopus)。这是首次在南半球发现的厚头龙类恐龙。
在1993年,纽约州立大学斯通尼布鲁克分校与马达加斯加的安塔纳那利佛大学展开了马哈赞加盆地计划,挖掘并研究马哈赞加省贝立佛查村附近的晚白垩纪地层中发现的化石与地质第一次的挖掘活动发现了数百颗兽脚类恐龙的牙齿,经鉴定后属于玛君龙,有些牙齿附着在一个前上颌骨之上,该前上颌骨在1996年被研究[16]。而接下来的七次挖掘活动发现了数万件的化石,其中许多化石属于新的物种。马哈赞加盆地计划在该地区发现了原有已知化石物种的五倍。
在1996年,这个挖掘团队发现了一个保存极良好的完整兽脚类头颅骨(FMNH PR 2100标本)。头颅的顶部有个半球形隆起物,类似苏伊士与塔丘特所叙述的玛君颅龙。在1998年,玛君颅龙再度被归类为阿贝力龙科恐龙,而非厚头龙类。虽然玛君龙(Majungasaurus crenatissimus)的命名比玛君颅龙(Majungatholus atopus)还早,但研究人员认为玛君龙模式标本的齿骨过于零碎,而没有足够的把握将它与玛君颅龙的头颅骨分类于同一物种。
在接下来10年,发现了一系列较不完整的头颅骨,以及数十个不同个体的部份身体骨骼,这些骨骸有幼年体也有成年体。该计划的成员也发现了数百件玛君龙的相关骨头,以及数千颗零散的牙齿。将这些化石组合过后,几乎拼凑出完整的骨骸,但大部分前肢、大部分的骨盆、以及尾巴的末端等部份仍未发现。这个挖掘计划在2007年达到顶峰,一份由7个关于玛君龙的科学研究,所集合的专题论文,在古脊椎动物化石学会的学会会议上发表。这个专题论文是以英文写成,但每份研究都有以马拉加斯语写成的概要。在论文之中,Lavocat将之前提到的齿骨重新鉴定,并将它分类于玛君龙。因此,玛君颅龙被玛君龙所取代。虽然该份专题论文内容非常广泛,但该论文只研究从1993年到2001年期间所发现的化石。而在2003年与2005年,在该地又发现了相当数量的标本,有些标本非常完整,这些化石正等待者更进一步的叙述